Haema 2019; 10(2):68-112
Konstantinos Liapis
Consultant Haematologist, Georgios Gennimatas Hospital
Full PDF | ![]()
Blood is examined for the following parasites:
– Plasmodium
– microfilariae
– Trypanosoma
-Leishmania
Νote: in some areas, malaria, microfilariae and/or trypanosomes may all be present and, therefore, it is not unlikely to detect two parasites e.g. Loa loa and P. falciparum in the blood film of a patient from Nigeria or Cameroon.
Blood films are critical in the diagnosis of malaria, filariasis, and trypanosomiasis. Bone marrow aspiration is important in the diagnosis of leishmaniasis and occasionally leads to the diagnosis of other parasitic infections. In rare cases, leishmania organisms may be seen in the granulocytes or monocytes of the peripheral blood.
Τhin films or thick films?
Either thin films or thick films may be used depending on the circumstances. Τhe thick film is more sensitive in detecting plasmodia and trypanosomes. Examine the centre portion of the thick film first (where it is more likely to detect parasites), and then examine the thinner outer edges (where the morphology is better). In thick films, organisms tend to be more compact and denser than in thin films (they often appear smaller). In contrast, the thin film causes very little distortion of the parasite, and permits species identification when it may not be possible in thick films.
The morphological features of malaria parasites may not be distinct in the thick film and, therefore, the thin film should be used for species identification.
Note:
– in most cases of malaria, the thin film will be positive. In order to increase sensitivity, look carefully in the thicker areas of the thin film (a ‘form’ of thick film).
– 3 consecutive negative thin films practically rule out malaria (repeat every 12-24 hours up to three; attempt to obtain blood samples during fever)
– if P. falciparum is seen, you should request a thick film for calculation of parasitaemia (parasites per μl οf blood).
– thick films should be used when searching for microfilariae (unlike Plasmodium and Trypanosoma, thick films cause no distortion of the morphological chracteristics of microfilariae).
1.1 MALARIA PARASITES1-12
See – Cases illustrating malaria parasites in: Illustrations of Blood Parasites (Figures 28-38)
See – Life cycle, epidemiology, and clinical information in Supplementary Appendix 1
Diagnosis of malaria is based primarily on the recognition of parasites in well stained thick and thin blood films stained with a Romanowsky stain (e.g. Giemsa, Leishman, Field) at pH 7.2-7.4. Thick films may improve sensitivity (it is said that the limit of detection of plasmodia is 10 parasites per mL blood versus 200 parasites per mL blood for thin films) but, as mentioned, a thin film is preferable for species identification.
In examining thin blood films for malaria, you must look at the appearance of the red cells containing the parasites and the appearance of the parasite.
1. Infected red cells
– Look at the size of the infected red cells
– Is the red blood cell filled with pink or red staining dots (Schüffner’s dots)?

* Not always! P. vivax or Ρ. ovale parasitised red cells may be normal sized, especially when they contain young trophozoites (rings).
** Schüffner’s dots are called James’ dots in the case of Ρ. ovale.
*** Schüffner’s dots may not be present in the early rings of P. vivax or Ρ. ovale. Also, in poorly stained slides, Schüffner’s dots may not be visible so it is essential that correct staining methods are used. In particular, the pH of the buffered saline used in the Giemsa stain has a profound effect on the staining of Schüffner’s dots and parasites.
Schüffner’s stippling versus basophilic stippling — what is the difference? 1. Schüffner’s dots are pink or red staining dots; basophilic stippling is blue or blue-purple 2. Unlike basophilic stippling, Schüffner’s dots will not be present in red cell without parasites.
Note: the appearance of Schüffner’s dots depends on the quality of the Giemsa stain.
The pH of the buffered saline used in the Giemsa stain is usually 6.8. The desired standard for malaria staining is 7.2. Schüffner’s dots may not be visible in slides stained with Giemsa at pH <6.8 (Figure 1).
Red cell stippling does not occur in P. malariae. However, fine pink-staining dots (called Ziemann’s dots) may appear in alkaline pH ≥7.5 or after prolonged staining. Ziemann’s dots do not appear if correct staining methods are used.
Tip: a simple, practical way to evaluate pH are the eosinophilic granules which are excellent pH markers because they stain bright red when the reagents are acidic and blue or dark grey when they are alkaline. Also, erythrocytes stain orange in acidic pH <6.4 and green in alkaline pH >7.6. In properly stained slides, eosinophilic granules stain red-orange and erythrocytes reddish or red.
Reliable and accurate parasite identification depends on good quality Giemsa.
Poor quality Giemsa may lead to technical errors.
Be sure to use newly prepared Giemsa. Don’t use Giemsa of dubious pedigree!
Parasites
Identification of malaria parasites
Things that may be confused with malaria parasites in blood films include platelets adhering to red blood cells, superimposed platelets Cabot’s rings, Howell-Jolly bodies, precipitated stain on red blood cells, unfiltered or contaminated Giemsa, debris from the patient’s skin, dust that gets on the blood film while it is drying and other artefacts that may appear on the slide during preparation. All these may look like malaria parasites in stained blood films and can be mistakenly identified as malaria. Usually, however, they do not have all three components of a malaria organism (i.e. the basic structure of malaria parasites).
The three components of malaria parasites are:
– blue staining cytoplasm
– red or purple staining chromatin
– brown or black pigment (haemozoin).
Except for early (young) rings (which lack pigment), you should see all three characteristics before you identify any structure as a malaria parasite. If you are uncertain whether a particular object or structure is a malaria parasite, look for more typical forms.
Table 1 shows the characteristics of the four main plasmodium species.

Figure 1. Effect of pH on Giemsa staining of malaria parasites.
Figures 2-5 are modified from: Βench aids for the diagnosis of malaria, WHO, 1988. Also, see selected cases illustrating malaria parasites in: Illustrations of Blood Parasites (Figures 28-38).
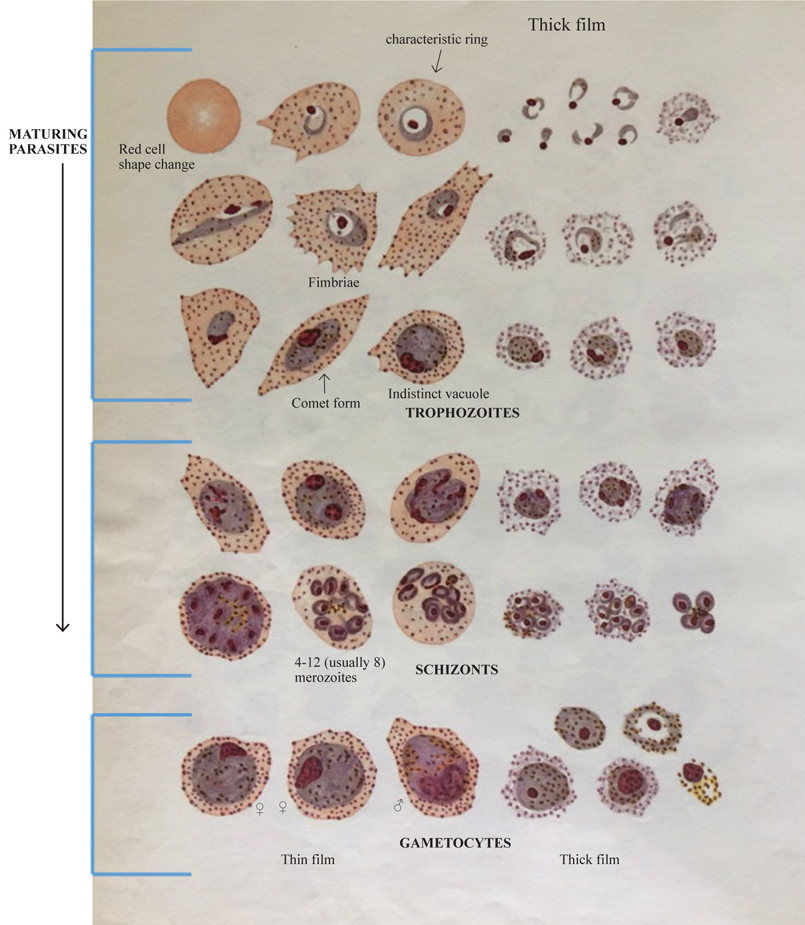
Figure 2. Plasmodium ovale (parasitaemia is often not high).
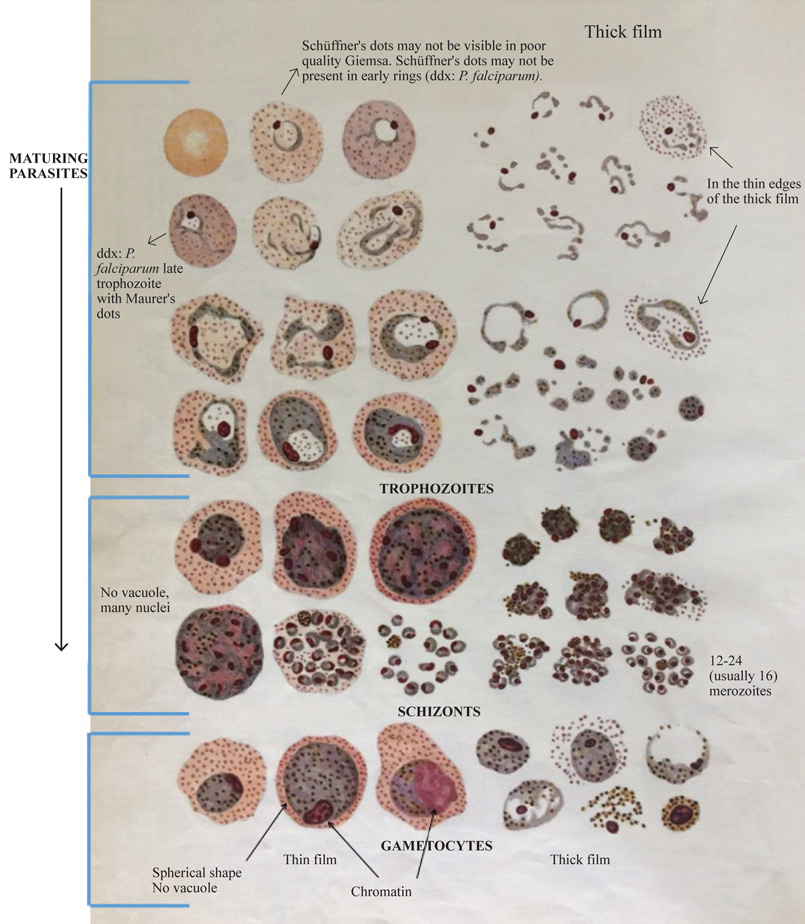
Figure 3. Plasmodium vivax.
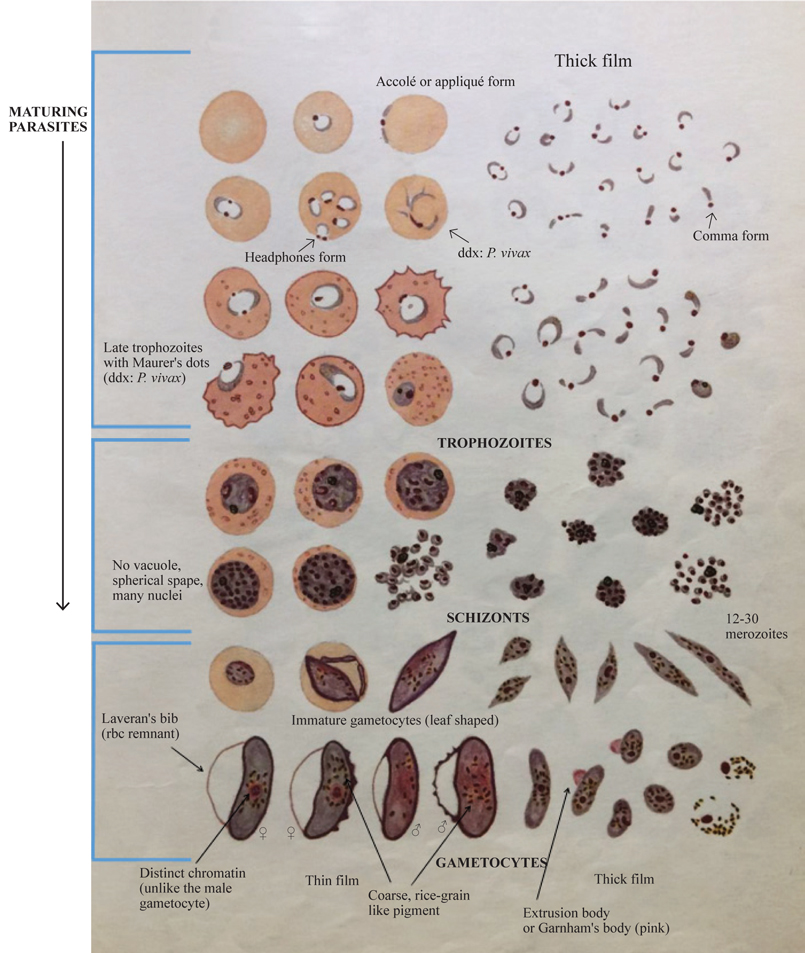
Figure 4. Plasmodium falciparum.

Figure 5. Plasmodium malariae (it is associated with low-level parasitaemia ≤1%).
Maurer’s dots versus Schüffner’s dots — what is the difference? Maurer’s dots are bigger, darker, and far less numerous than Schüffner’s dots (the red cell is not entirely filled in contrast to Schüffner’s stippling).
Rings of the four species may look alike. If you see rings, look for older stages (late trophozoites, schizonts, and gametocytes) → older stages should help you identify the Plasmodium species.
In patients with P. falciparum only rings are usually seen (or rings + gametocytes or only gametocytes); older stages (schizonts) are present only in severe infections. Occasionally, late trophozoites may be seen.
P. falciparum schizonts are rare in peripheral blood films (only in severe infections with high parasitaemia) because schizonts sequester in the microcirculation (through small protuberances or knobs of the erythrocytic membrane) causing obstruction. Pfemp-1, an important cell adhesion molecule on knobs, interacts with ICAM-1 on endothelial cells (→ sequestration) and with CD36 on adjacent red cells (→ rοsetting).
In contrast, late trophozoites and mature schizonts are common in blood films of patients with other malaria species.
How to identify trophozoites, schizonts, and gametocytes:
Schizont → a schizont contains ≥2 nuclei (because schizogony has occured)
Note: the nucleus of P. falciparum or P. vivax rings may include two chromatin dots (like ‘headphones’).
Trophozoite → contains 1 nucleus and vacuole (usually)
Gametocyte → contains 1 nucleus without vacuole
P. falciparum → (usually) rings and/or gametocytes present in peripheral blood (rarely: late trophozoites, schizonts)
P. falciparum gametocytes appear in the peripheral blood after approximately one week of fever.
P. vivax → (usually) all stages present in the peripheral blood.
If P. falciparum is seen, the level of parasitaemia must be reported.
You can estimate the level of parasitaemia by three methods:
- % parasitaemia (number of infected red cells per hundred)
Using 100× (oil immersion) objective, count the number of infected erythrocytes in 5 fields of a thin film (~1000 erythrocytes).
- parasitaemia/μl (parasites per microlitre)
The following is a practical method of adequate accuracy. This method is based on the number of parasites per μl of blood in a thick film, these being counted in relation to a predetermined number of leukocytes. Two tally counters are required to count parasites and leukocytes separately in a thick film. Using 100× (oil immersion), count the number of parasites (P) and leukocytes until 300 leukocytes have been enumerated.
Parasitaemia (/μl) = WΒC × P/300
(WΒC = white blood cell count)
Count all the species present (some cases are mixed infections) and include both sexual (gametocytes) and asexual forms together.
This method must always be used in cases of suspected drug-resistant parasites.
- The plus system using the 100× (oil immersion) objective in a thick film:
+ = 1-10 parasites/100 fields
++ = 11-100 parasites/100 fields
+++ = 1-10 parasites/1 field
++++ = >10 parasites/1 field
The plus system is not very accurate and should be used only when it is not possible to undertake the more acceptable parasite count per μl of blood.
The parasitaemia/μl method and the plus system are based on the estimation that about 100 fields, using a 100× (oil immersion) objective, is the equivalent of 0.25 μl of blood.
Haematological changes associated with malaria:
Malaria may be accompanied by atypical (reactive) lymphocytes in the blood film (usually <15% of the leukocyte count ≠ infectious mononucleosis). It is not uncommon to find plasmacytoid lymphocytes (plymphs) and Türk cells and you may even find circulating immunoblasts (activated large Β-cells) with deeply staining basophilic cytoplasm, high nuclear:cytoplasmic ratio, and nucleoli (Figure 6).
Note:
lymphoplasmacytoid cells may be seen in the peripheral blood in cases of immune system activation by various stimuli.
reactive lymphocytosis is more common and prominent in another tropical disease that is very similar to malaria, dengue, which is endemic in South-East Asia, western Pacific, and the Caribbean.

The neutrophil count is usually low or low-normal (due to margination to the endothelial surface), normal, or slightly increased. The platelet count is often reduced in acute malaria (the mean platelet count is 132×109/l at presentation and rises to 234×109/l, 12 days after treatment), however, severe thrombocytopenia (<40×109/l) is rare (5%). Several mechanisms are responsible for the thrombocytopenia: splenic pooling, platelet activation, platelet binding to CD36 on infected red cells, and antibody mediated platelet destruction. Megakaryocytes in the marrow are usually normal or increased in number (i.e. peripheral thrombocytopenia).
Note: platelets are often abnormally large in malaria (e.g. MPV 10.5 fl) so that the automated platelet count may be falsely low.
A major mechanism of haemolysis in malaria is the rupture of red cells at the time of release of merozoites (i.e. haemolysis in malaria is linked to the red cell infection and is related to the level of parasitaemia). Coombs’ test is usually (-). However, P. vivax may rarely cause Coombs (+) warm autoimmune haemolytic anaemia with spherocytosis, and some antimalarial drugs can cause haemolytic anaemia in individuals with G6PD deficiency. Artemisin, the drug of choice for severe malaria, may rarely cause severe intravascular Coombs (-) delayed haemolytic anaemia, occurring 7-31 days after treatment. The mechanism is unknown.
During acute malaria there is immobilisation of iron in the macrophages, and low serum iron concentrations: serum ferritin is increased (acute reactive protein). Serum folic acid is normal or low. Red cell folate levels are raised above normal.
- P. falciparum and P. knowlesi can lead to severe, life-threatening malaria with organ failure. P. vivax is less commonly associated with severe malaria than P. falciparum.
The morphological clues to severe P. falciparum malaria include (see Figure 7):
– parasitaemia ≥5% (‘hyperparasitaemia’)
– neutrophil leukocytosis >15×109/l and toxic granulation (secondary bacterial infection?)*
– schizonts present in the blood film
– malaria pigment in neutrophils and monocytes (pigmented leukocytes may also be seen in cases with delayed diagnosis or in recurrent malaria)
– features of hyposplenism (e.g. Howell-Jolly bodies) (post-splenectomy or asplenic patients run a significantly increased risk of severe malaria).
* Malaria is immunosuppressive and is frequently complicated by secondary infections such as bronchopneumonia, urinary tract infections, and Gram-negative septicaemias.
Note: high-density parasitaemia may lead to severe intravascular haemolysis with haemoglobinuria (‘blackwater fever’). More commonly, however, haemoglobinuria is due to haemolysis developing in G6PD-deficient patients in response to oxidant antimalarials e.g. primaquine or quinine.

Figure 7. A and B. Overwhelming P. falciparum infection in the ICU (the patient was a pilot who returned from Ivory Coast 5 days before admission to ICU): parasitaemia 6%, pigmented neutrophil with 4 lumps of haemozoin (>>), Howell-Jolly body (arrow), schizont (*), echinocytes due to acute renal failure, thrombocytopenia, and red cell fragments (+) due to DIC.
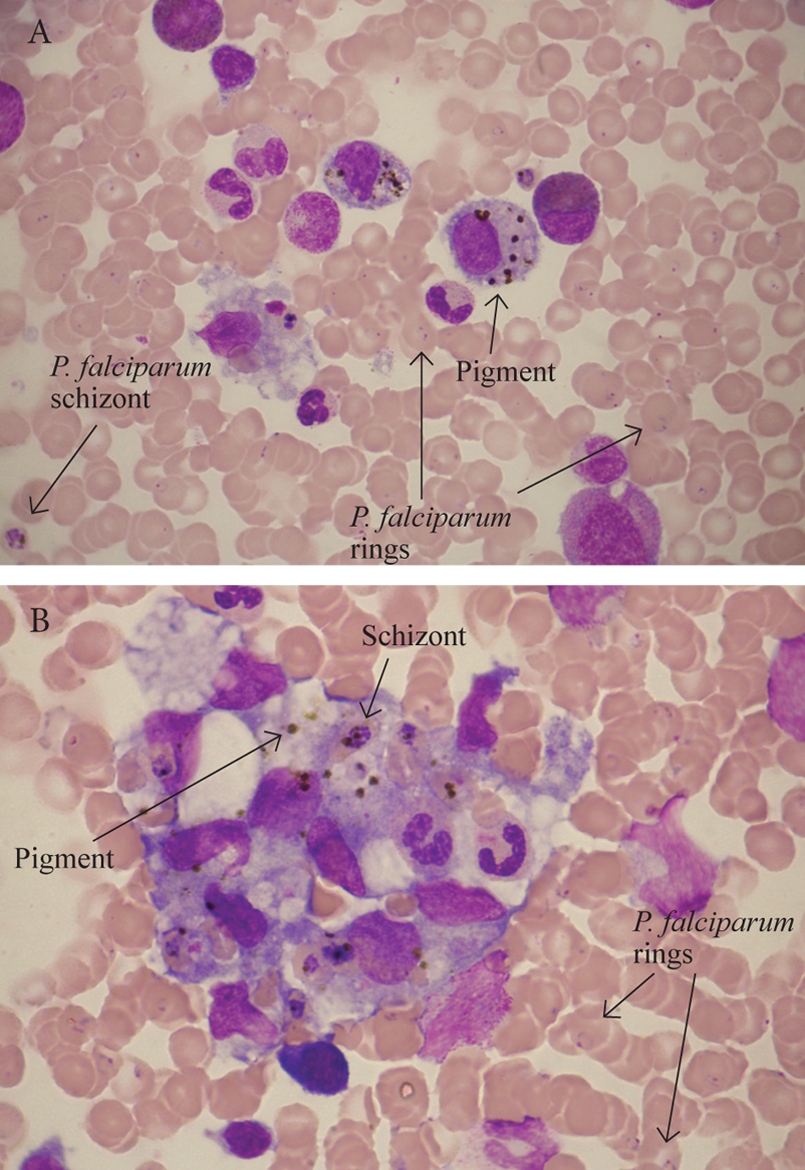
Figure 8. A and B. Bone marrow aspiration of a patient with heavy P. falciparum parasitaemia, pancytopenia, and haemophagocytosis. Malaria pigment is crystalline and birefringent and, thus, easily seen with polarised light.
Patients with few parasites detectable on initial blood film may in fact be seriously ill due to parasitised red cells being sequestered in internal organs. Therefore, the intensity of the parasitaemia in the peripheral blood does not always mirror the intensity of the infection.
Bone marrow aspiration is rarely performed in malaria (e.g. for investigation of excess cytopenia). Bone marrow findings include:
– hypercellularity with increased macrophage activity
– phagocytosed parasites and/or pigment in bone marrow macrophages (haemozoin may also be seen within neutrophils and granulocytic precursors)
– dyserythropoiesis (binucleated or multinucleated normoblasts, poor haemoglobinisation, basophilic stippling, intracytoplasmic bridges, nuclear abnormalities e.g. nuclear budding, irregularity, and karyorrhexis) which leads to ineffective erythropoiesis. The mechanism of dyserythropoiesis is uncertain, but it could be secondary to hypoxia from the packing of bone marrow sinusoids with parasitised red cells, or be mediated by an imbalance of cytokines (malaria is associated with high levels of IFNγ and TNFα which may lead to activation of the extrinsic pathway of apoptosis in erythroid cells).
– megaloblastic change e.g. giant metamyelocytes
– haemophagocytosis and, rarely, haemophagocytic syndrome (Figure 8)
– P. falciparum schizonts and immature gametocytes (not usually seen in the peripheral blood)
– toxic effects of antimalarial drugs (e.g. megaloblastic change or neutropenia with a maturation block)
– rarely, malaria may be diagnosed from a bone marrow aspirate smear!
Note: some antimalarials e.g. malarone (proguanil/atovaquone) may cause severe acute megaloblastic anaemia with pancytopenia in patients with chronic renal failure even in prophylactic doses.
About 0.5-1% of malaria cases are mixed infections e.g. P. falciparum + P. ovale, as shown in Table 2. These cases are very difficult (your worst nightmare!). The malaria rapid diagnostic test (RDT), a chromatographic test, may help to diagnose these cases: it detects separately P. falciparum Ag (PfHRP-2), P. vivax Ag (P. vivax pLDH), and a panmalarial Ag (Plasmodium aldolase) in blood.
The level of parasitaemia is useful in the differentiation of the different Plasmodium species
In P. falciparum, it can be very high e.g. parasitaemia 12% is not uncommon in severe P. falciparum infection with severe anaemia (levels >20% are rare even in high grade infections). P. vivax and P. ovale are associated with a level of parasitaemia up to 2-3%. P. malariae causes a low grade infection with less than 1% parasitised red cells which highlights the importance of careful examination of the blood film in suspected malaria. Since P. malariae parasitaemia is max 1%, the development of anaemia takes a long time.
The travel and migration history may also aid in the differentiation of malaria parasites
In patients who return or come from:
– Tropical Αfrica → consider P. falciparum
– North Africa → consider P. vivax
– West Αfrica → consider P. falciparum, P. ovale
– Middle East (e.g. Turkey, Pakistan, Afghanistan) → consider P. vivax
– Central America (e.g. Μexico) → consider P. vivax
– Haiti → consider P. falciparum
– South America, India → P. vivax ~ P. falciparum
– South-East Asia → P. falciparum ~ P. vivax (consider P. knowlesi too)
– Malaria in migrants who live exclusively in non-endemic regions for a long time → consider P. vivax or P. ovale relapse
– Accutely ill patient requiring admission to ICU → consider P. falciparum (rarely P. vivax)
Ιt is important to re-examine the blood film after treatment to rule out resistance in falciparum malaria (rings should have disappeared in the peripheral blood on day 2 and gametocytes on day 6).
Request a thick film on:
– Day 2 (in severely ill patients) and day 4 (in less sick patients): if parasitaemia/μl (rings+gametocytes) is 20-25% above the level before treatment or if the ring forms are reduced by less than 75% → it indicates a resistant strain (high level of resistance, R-III level) and a need for a change in treatment.
– Day 7: no parasites (rings+gametocytes) should be present. A film should be examined on day 28 to rule out a strain with late recrudescence (low level of resistance, R-Ι level).
– If the ring forms have disappeared on day 4 but return and are present on day 7 → the parasites are resistant (R-Ι level)
– If the ring forms are still present on day 2 but the parasite count is reduced by ≥75% → it indicates an intermediate level of resistance (R-II level).
Drug resistance is seen in P. falciparum and rarely in P. vivax.
Following clearance of the parasitaemia and during recovery, the blood film shows a reticulocytosis, anisocytosis, macrocytosis, and polychromasia. Leukocytes containing malarial pigment can be found in the blood for many days after parasitised red cells have disappeared; this can be useful in making a retrospective diagnosis of malaria.
Morphological differential diagnosis of malaria:
– The gametocytes of P. falciparum may be confused with Trypanosoma (the presence of pigment helps to differentiate between them).
– Bizarre forms can occur: e.g. P. vivax ookinete, which is crescent-shaped, looks like P. falciparum gametocyte. Ookinetes are formed inside the midgut of the mosquito host, but they can rarely be found in the blood film (their presence indicates a substantial delay has occurred between the time the blood was collected and the time the film was prepared). P. vivax ookinete has pigment in apical position and usually a small vacuole ≠ P. falciparum gametocyte.
– You may also see in the blood film the exflagellated form of male microgametocyte which is normally formed inside the mosquito host (Figure 9). It indicates a substantial delay has occurred between the time the blood was collected and the time the film was prepared. Microgametocyte exflagellation has been observed in all four Plasmodium species. The importance of exflagellated microgametocytes is that they can be misdiagnosed as trypanosomes or relapsing fever Borrelia spirochaetes (relapsing fever is a tropical disease that manifests with recurrent fevers, flu-like symptoms and splenomegaly like malaria).
Crescent shaped forms in an otherwise typical case of P. vivax → consider P. falciparum co-infection or ookinete formation
Blood films should be prepared no longer than 3-4 hours after blood collection to minimise the risk of distorted morphology and the potential appearance of parasite stages not normally occurring in the blood.
– The differential diagnosis (ddx) of P. falciparum malaria also includes babesiosis, caused by small blood protozoan parasites called piroplasms.
Babesiosis (piroplasmosis)6-12
The geographical distribution of babesiosis is different from malaria. Babesiosis is not endemic in the tropics but in the north temperate zones. Most human cases of babesiosis are due to two species:
– Babesia microti: in northeastern coastal areas of USA e.g. New England (Massachusetts, Rhode Island, Connecticut, Nantucket Island) and New York State, typically occurring between May and October. B. microti is a parasite of rodents (white-footed mouse). In the Washington State and California, babesiosis is caused by another small Babesia species, B. duncani, also known as WA1 (Washington 1) and CA1-CA4 (California 1-4). B. microti and B. duncani are morphologically indistinguishable.
– Babesia divergens: in Europe (e.g. France, Austria, England, Scotland, Ireland, Romania). B. divergens is a major parasite of cattle (very important to veterinarians).

Figure 9. Exflagellated microgametocyte of P. vivax.
Human babesiosis is a zoonosis transmitted by the bite of hard ticks (family Ixodidae) which parasitise a wide range of wild and domestic hosts: mice, deer, and cattle (patients often have a history of rural work or camping).
Babesiosis involves individuals who have had splenectomy (Howell-Jolly bodies+), in whom it can cause extreme parasitaemia (up to 85%), fulminant intravascular haemolysis, haemoglobinuria, exhaustion, and death within a week. In those with an intact spleen, the severity of the infection varies from a subclinical or mild infection to an acute haemolytic anaemia requiring hospitalisation (parasitaemia 1-10%).
Risk factors for more severe babesiosis include age >40 years, asplenia, and immunocompromised patients such as those with HIV, corticosteroids, or other immunosuppressive therapy.
Babesiosis is an emerging problem in transfusion medicine in USA: transfusion-transmitted infection, like naturally occurring infection, is more often recognised in asplenic or hyposplenic patients including patients with sickle cell disease.
Babesiosis is morphologically very similar to P. falciparum malaria: in both conditions, red cells contain ring trophozoites (frequently with a clear central vacuole), often 2 or 3 parasites per rbc (multiple rings), one or two chromatin dots, appliqué or accolé forms at the edge of the rbc may be present, and parasitaemia may be high. Thus, Babesia may easily be confused with P. falciparum.
Β. microti ring forms may look like P. falciparum rings. However, piroplasms are smaller (1.2-2.0 μm), often appear as small coccoid parasites, have no pigment, and there are no Schüffner’s or Maurer’s dots, schizonts, or gametocytes. Babesia multiplies in the red cell by budding (whereas Plasmodium multiplies by schizogony), and one organism forms up to four merozoites. This process results in a limited variety of small coccoid, pyriform or ring forms (single rings, paired rings, and tetrads so-called ‘Maltese cross’ formations, which although rare, are essentially pathognomonic of babesiosis since they are not seen in malaria). Extracellular parasites (merozoites) may be seen in the peripheral blood film and can form clusters.
Β. divergens is similar in size to Β. microti (1.0-2.4 μm) but more frequently pyriform in shape (pear shaped), which is typical of a piroplasm (piro = pear in Latin). B. divergens takes the form of pear shaped, oval or round ring forms, multiple ring forms, coccoid and amoeboid shapes, and may exist in pyriform pairs (paired forms, diverging at a wide angle of up to 180°, typically situated on the periphery of the red cell known as divergent forms) and tetrads (Maltese cross formations). A predominance of pyriform pairs (divergent forms) is characteristic
Β. divergens. There may be one to eight parasites per rbc. B. divergens is often more severe than B. microti.
Note: Babesiosis is often associated with lymphopenia and thrombocytopenia. Lymphocytosis with atypical lymphocytes may also occur in babesiosis, as in malaria.
Post-babesiosis warm autoimmune haemolytic anaemia with erythrophagocytosis by neutrophils in the blood film may occur 2-4 weeks later. Haemophagocytosis has been observed in the bone marrow in babesiosis and rarely frank haemophagocytic syndrome may develop.
Tips:
Co-infection with Lyme disease should be ruled out in any case of babesiosis (same vector: ixodes ticks). Diagnosis: Lyme disease IgM and IgG Ab.
Malaria RDT is negative in babesiosis.
Babesiosis is a serious infection of cattle. Figure 10 shows the main species of Babesia (B. bigemina, B. bovis, and B. divergens) in bovine erythrocytes. Most human cases are due to B. divergens; rarely, B. bigemina and B. bovis have been found to infect humans.
1.2 ΤRYPANOSOMA3-12
> See – Cases illustrating the microscopical features of African trypanosomiasis in: Illustrations of Blood Parasites (Figures 39 and 40)
> See – Life cycle, epidemiology, and clinical information in Supplementary Appendix 2
The prime means of diagnosing African trypanosomiasis or African sleeping sickness is based on the observation of the flagellated protozoan Trypanosoma brucei in the blood film. There are two subspecies of Trypanosoma brucei: Τ. b. gambiense in western and central Africa (‘Gambian’ trypanosomiasis) and T. b. rhodesiense in eastern and central-eastern Africa (‘Rhodesian’ trypanosomiasis). Trypanosomes can be identified (depending on the stage of the disease) in:
– aspirates from the trypanosomal chancre
– blood films
– lymph node aspirates
– bone marrow aspirates
– cerebrospinal fluid (CSF) cytospins

Figure 10. A. Main species of Babesia in bovine erythrocytes. B. Morphological features of B. dirergens.
Only the trypomastigote form of T. brucei is found in humans. Like plasmodia, trypanosomes may be very difficult to detect in the blood film, especially Τ. gambiense in which parasitaemia is often very low (≠ T. rhodesiense detection is much easier). The examination of thick films, buffy coat films (prepared by the microhaematocrit method), and concentration methods (mini anion exchange centrifugation) can increase the chances of detection. In the thick film, trypanosomes may be distorted and difficult to differentiate from cellular debris.
Delays in diagnosis are common e.g. a case of Τ. b. gambiense with a long delay in diagnosis has been reported despite multiple admissions and lymph node biopsy in which the diagnosis was finally made by bone marrow aspiration 6 years after exposure.
Identification of African and American trypanosomes in blood films (Figure 11)
Trypanosomes are more readily identified in the thicker areas of the thin blood film (they are between the red blood cells). Look for an elongated organism with a nucleus located near the centre, a kinetoplast located near one end and a free flagellum extending from the other end. Look at the length, the shape, and the size of the kinetoplast of the parasites.
In Africa, the two T. brucei subspecies that infect man are morphologically identical. You cannot determine the subspecies from the appearance in stained film; however, epidemiology and clinical picture are different:
T. b. rhodesiense → causes more acute disease (high level parasitaemia)
T. b. gambiense → causes more chronic disease (parasitaemia may be low)
In South and Central America, T. cruzi and T. rangeli may rarely be found in blood films. T. cruzi causes American trypanosomiasis (Chagas’ disease), whereas T. rangeli, although it may circulate in blood, appears to be non-pathogenic to man and its only medical importance lies in the possibility of its being mistakenly diagnosed as T. cruzi. T. cruzi and T. rangeli can be morphologically distinguished from each other:
T. rangeli is longer and thinner (long, slender trypanosome) than T. cruzi, the nucleus is in more anterior position (central in T. cruzi), its kinetoplast is small (similar to African trypanosomes), and is often seen in S shape.
T. cruzi has a very large, prominent kinetoplast and is often seen in C, U, or (less commonly) S shapes.
T. cruzi is badly distorted in thick films and is more readily identified in the thicker areas of the thin film.You can find it in:
– its trypomastigote form in peripheral blood films during acute Chagas’ disease (high level parasitaemia); usually it disappears from blood films during the chronic phase of Chagas’ disease (low level parasitaemia).
– its trypomastigote form or its amastigote form (leishmanial form) either extracellularly or inside macrophages in bone marrow aspirates, following reactivation of Chagas᾿ disease in immunocompromised hosts. The leishmanial form of T. cruzi should be distinguished from L. chagasi. HIV(+) patients with T. cruzi reactivation typically present with meningoencephalitis or cerebral tumour-like lesions ± myocarditis (ddx: Toxoplasma).
Note: reactivation of Chagas’ disease in immunocompromised patients may manifest as acute Chagas’ disease with fever and high-level parasitaemia → you may find trypanosomes in peripheral blood films or in CSF preparations.
Like plasmodia, the cytoplasm of Τ. brucei stains blue. The nucleus (located near the centre of the body) and kinetoplast stains red or purple. The kinetoplast, seen as a small dot, is located near one end of the organism (posterior end) and the free flagellum extends from the other end (anterior flagellum). The flagellum originates from the posterior part of the trypanosome next to the kinetoplast. The flagellum is attached to the cell membrane except at the anterior tip, where it terminates with a free end. Trypanosoma is a motile organism: since the flagellum is constanty moving it pulls the cell membrane into irregular extensions, this feature being known as the undulating membrane. The trypanosome may be undulated with 2 (S shape) or 3 curves or it may be in a C or U shape.

Figure 11. Trypomastigotes in blood films.
The length of T. cruzi is 20-25 μm; T. brucei is 13-30 μm (usually 20-29 μm); T. rangeli is 30 μm. The undulating membrane is difficult to see in T. cruzi. T. cruzi kinetoplast is larger and thicker than Τ. brucei kinetoplast. T. cruzi nucleus is located at the centre of the body; Τ. brucei nucleus is located near the centre of the body. The cytoplasm may contain irregular-sized bluish granules.
The trypanosomes have one large mitochondrium. The kinetoplast is the mitochondrial DNA. The flagellum originates from the parabasal body close to the kinetoplast, and it is not connected with the kinetoplast (Figure 12).

Figure 12. Ultrastructure of T. b. gambiense.
Tips:
– African trypanosomes may be numerous (especially T. rodensiense) or difficult to find. Look for elongated organisms with the characteristic basic structure: central nucleus – small kinetoplast – flagellum.
– with careful observation, you will see slender, stumpy, and intermediate forms of T. brucei. The slender trypanosomes are the diving forms. The stumpy forms are not diving. In the tsetse fly vector, stumpy forms transform to dividing epimastigote forms.
– when the trypanosomes have passed the blood-brain barrier and invade the CNS, the patient is in the second phase of the disease. The only way to diagnose trypanosomes in the nervous system is to examine the CSF for trypanosomes. The presence of morula cells of Mott or an elevated CSF IgM is thought to strongly suggest CNS involvement. An elevated CSF cell count usually accompanies these findings. – Trypanosoma is interesting to the haematologist for one more reason: the surface coat of T. brucei consists of a dense layer of a single protein species, the variant surface glycoprotein (VSG). T. brucei changes its VSG coat regularly in order to evade the immune defense of their mammalian host (i.e. antigenic variation). VSGs were the first GPI-anchored proteins to be discovered in 1985, leading to the discovery of GPIs in ΡΝΗ in 1986.
1.3 ΜICROFILARIAE3-13
> See – Cases illustrating microfilariae in: Illustrations of Blood Parasites (Figures 41-44)
> See – Life cycle, epidemiology, and clinical information in Supplementary Appendix 3
Microfilariae are bloodstream threadlike nematode helminths. The following characteristics are used for the identification of microfilariae:
– size of the microfilaria
– row of nuclei (dispersed or crowded)
– presence or absence of a sheath
– presence or absence of nuclei in the tip of the tail
– head space (long or short)
– innerbody (can or cannot be demonstrated)
Identification of microfilariae
Fibres, fibrin, threads, hair, and other artefacts found on blood films are often confused with microfilariae especially if the patient has eosinophilia — the absence of nuclei rules out identification of these structures as microfilariae. Microfilariae have the characteristic basic structure: sheath—head—body—tail; they do not contain vacuoles and they are not refractile or septated.
Note: spores of helicosporous fungi (e.g. Ηelicosporium, Helicomyces) that get on the blood film while it is drying, may be mistaken for microfilaria (Figure 13).

Figure 13. Spores of helicosporous fungi may be mistaken for microfilariae.
Three microscopical methods are used for detection of microfilariae in peripheral blood:
Thick films>Thin film capillary blood>Thin film venous blood
(Gold standard → thick film capillary blood)
Use a 10× objective to locate microfilariae (search the entire blood film systematically) and switch to 40× and 100× (oil immersion) objectives to examine microfilariae for specific identification. Also, remember to look for malaria parasites with the 100× objective.
Unfortunately, not all the diagnostic features can be seen in Giemsa stained films, so occasionally special stains such as Delafield’s haematoxylin or diluted haematoxylin must be used to demonstrate them.
A concentration method may be used to increase sensitivity (dilute blood 1:2 in distilled water or saponin saline solution or 2% formalin to lyse red cells, spin at high speed and examine deposit).
Microfilariae seen in blood films include:
– Wuchereria bancrofti → tropical Africa, Αsia, Egypt, Central and South America, Caribbean, Indian Ocean
– Wuchereria bancrofti var. pacifica → Pacific Ocean (Melanesia, Polynesia, Caledonia)
– Brugia malayi → southern India, South-East Asia e.g. Thailand and the Philippines. (southern China and Korea are now free of infection)
– Brugia timori → South-East Asia (Indonesian archipelago, Timor Island)
– Loa loa → West and Central Africa
– Mansonella perstans → tropical Africa
– Mansonella ozzardi → Latin American and Caribbean region (infects humans but is probably non pathogenic)
The most important filariases are W. bancrofti and B. malayi (→ lymphatic filariasis: 90% Wuchereria, 10% Brugia) and Loa loa (→ loiasis).
In cases of suspected filariasis*, the time of day is important in taking blood samples for examination because some species have a periodicity. That is, microfilariae, released from the uterus of adult female worms, are present in the blood only at certain times of the day and in order to detect their presence, the blood must be collected at the appropriate time (῾window᾿ of detection), as shown in Table 3.
* Filariasis is usually suspected in travellers, migrants, or servicemen with eosinophilia ± fever returning from the tropics. Other signs suspicious for filariasis include high IgE, hydrocele, lymphadenitis/lymphadenopathy (inguinal, axillary or generalised), lymphagitis, and lymphoedema (lymphatic filariasis). Note: Loa loa is usually accompanied by eosinophilia and very high IgE but eosinophilia may be absent from some patients with lymphatic filariasis.
Microfilariae may also be observed in lymph node aspirates usually in a background of a mixed population of cells comprising of mature lymphocytes, centrocytes, centroblasts, histiocytes, and eosinophils.
Microfilariae may occasionally be observed in bone marrow aspirates, more commonly in immunocompromised hosts e.g. patients with HIV, haematological malignancies, and solid tumours. An increased number of bone marrow eosinophils and their precursors are typically seen. Bone marrow filariasis may be asymptomatic or lead to aplastic, hypoplastic, or hyperplastic marrow with a normoblastic or a megaloblastic appearance.
In some cases, involvement of the bone marrow is associated with marrow hypoplasia, fever, and pancytopenia which may be severe. Whether these cases represent co-existence of filariasis with aplastic anaemia or toxic suppression of the bone marrow mediated by liberation of toxic metabolites by growing larvae (microfilariae) or cytokines is unclear. The majority of reported cases of aplastic anaemia with microfilariae in the bone marrow smears were W. bancrofti infections in patients from India.
A typical microfilaria appears serpentine in shape and filled with the nuclei of many cells; there are clear spaces (anucleate) which correspond to anatomic landmarks (Figure 14).

Figure 14. Diagram of a typical microfilaria (W. bancrofti). 
Figure 15. Sheathed and unsheathed microfilariae.
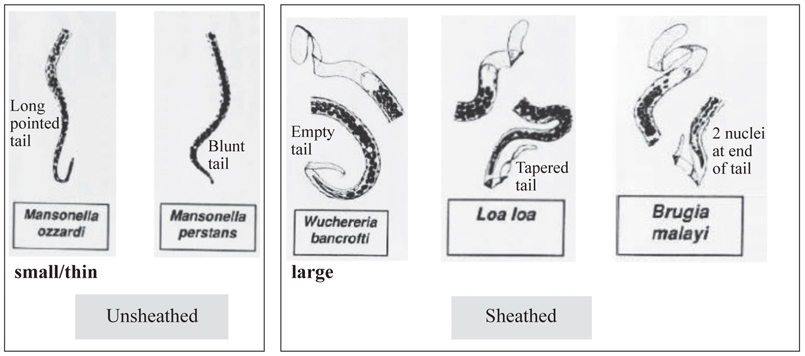
Figure 16. Key features of microfilariae.
The presence of a sheath is a major virulence factor for microfilariae (it inhibits the attraction-aggregation of eosinophils to the microfilaria; binds complement inhibitors factor H and C4bBP and blocks complement activation). Microfilariae without a sheath (M. perstanns and M. ozzardi) are doubtfully pathogenic microfilariae (Figure 15). A pathogenic microfilaria has a diametre of approximately as much as a lymphocyte (8-10 μm); the diametre of a non-pathogenic microfilaria is smaller (3-5 μm).
Morphological differentiation of microfilaria species
The presence or absence of a sheath and the presence or absence of nuclei in the tip of the tail are the two most important morphological features used for the identification of microfilariae (Figure 16). Sometimes the tip of tail may be folded beneath the body in the thin film. If you are uncertain, examine a thick film (it ῾unfolds᾿ the microfilariae).
Wuchereria bancrofti (pantropical: South-East Asia, India, Central Africa, Egypt, Central and South America, West Indies) and Wuchereria bancrofti var. pacifica (South Pacific Islands e.g. New Caledonia, Fiji, Samoa):
– large microfilaria 244-296 μm (mean, 260 μm); few, smooth body curves
– sheath may or may not stain pink (light) with Giemsa; does stain better with haematoxylin stains
– discrete nuclei (fewer body nuclei than other microfilariae); empty space between the nuclei and the body wall
– tail tapers but no nuclei in tip of tail i.e. empty tail sometimes the tip of tail may be bent back underneath the body
– innerboby may be visible in Giemsa (pink); does not stain with haematoxylin
– head space as long as it is broad (short head space)
Brugia malayi (South-East Asia, India):
– smaller than Wuchereria, 177-230 μm (mean, 220 μm)
– kinked, coiled microfilaria (many angular curves)
– sheath stains deep pink with Giemsa and is very prominent, does stain (light) with haematoxylin stains
– dense body nuclei (nuclei crowded and fill the whole body); empty space between nuclei and body wall
– tail tapers irregularly with 2 distinct nuclei at end of tail (subterminal and terminal nuclei widely separated)
– innerbody may or may not stain in Giemsa (when it does, it is prominent); does not stain with haematoxylin
– head space twice as long as it is broad (long head space)
Brugia timori (confined to part of Indonesia in South-East Asia):
– similar to B. malayi (long head space, widely separated subterminal and terminal nuclei) with 2 differences: larger in size 265-323 μm (mean, 287 μm) and sheath unstained in Giemsa
Loa loa (only in West and Central Africa):
– length 231-250 μm (mean, 238 μm)
– kinked (corkscrew appearance, several kinks and small curves)
– sheath does not stain with Giemsa (its outline may be demarcated by red blood cells); does stain with haematoxylin stains
– the column of nuclei is compact (nuclei crowded, almost indistinct); no empty space between nuclei and body wall
– tip of tail tapers, nuclei extending to tip of tail
– very frequently the tail is flexed or coiled within the sheath
– innerbody does not usually stain
– head space as long as it is broad (short head space)
Mansonella perstans (West and Central Africa and South America):
– small 190-200 μm (mean, 195 μm) and thin (4-5 μm)
– kinked
– does not have a sheath
– nuclei stain deeply and ‘run together’ (the column of nuclei appears more compact in Giemsa than in haematoxylin)
– nuclei extend to tip of tail; last nucleus is bigger; tip of tail is bluntly rounded (blunt tail)
– short head space
Mansonella ozzardi (Central and South America, especially Brazil, Guiana, Colombia, and the Caribbean):
– small 163-203 μm (mean, 183 μm) and thin (3-5 μm)
– stains very lightly
– kinked
– does not have a sheath
– the column of nuclei is compact
– nuclei do not extend to end of tail; tail tapers (tail is long, slender, devoid of nuclei and pointed)
– short head space (head space that is slightly longer than it is wide)
Note: It is not unsual to find two species of microfilaria in the same blood film:
– Wuchereria bancrofti + Brugia malayi in India and South-East Asia
– Mansonella perstans + Loa loa in Africa
– Mansonella ozzardi + W. bancrofti in South Africa
If M. perstans is seen, search for Loa loa! Also, look for malaria parasites (especially P. falciparum) with the oil-immersion objective. In Loa loa, consider ΗΙV-1 and HIV-2 testing (West and Central Africa).
Tips:
in cases of suspected filariasis (e.g. eosinophilia and leg oedema in a migrant) → request day and night blood films (capillary blood is preferred)
in general, microfilaraemia is higher in L. loa than in other microfilariae. However, amicrofilaraemic Loa loa infection does occur in people living in endemic regions so-called ‘occult loiasis’, even when high sensitivity methods are used (i.e. concentration techniques such as centrifugation and blood microfiltration). In such cases, diagnosis can be made by serology or by PCR (test of choice in amicrofilaraemic patients).
calculation of microfilaraemia is particularly important in Loa loa.
Microfilaraemia is calculated with the use of a thick film (as described for P. falciparum). This method is based on the estimation that about 100 fields, using a 100× (oil immersion) objective is the equivalent of 0.25 μl of blood. If Loa loa is seen, counts of microfilariae per unit quantity of blood are necessary before the administration of diethylcarbamazine (DEC) or ivermectin because high grade L. loa microfilaraemia (≥8000 microfilariae/ml) is associated with serious complications such as high fever, encephalopathy (headache and confusion, progressing to stupor and coma), glomerulonephritis, systemic allergic reaction (Mazzotti reaction) and excess mortality owing to the rapid death of microfilariae and massive degranulation of eosinophils causing inflammation.
Note: microfilariae and macrofilariae produce little reaction in the host while living but cause acute inflammation upon their death and liberation of their products or when they exsheath.
1.4 LEISHMANIA3-10,12,14
> See – Cases illustrating the microscopical features of leishmaniasis in: Illustrations of Blood Parasites (Figures 45-48)
> See – Life cycle, epidemiology, and clinical information in Supplementary Appendix 4
Leishmaniasis is caused by infection with the flagellate protozoan parasites of the genus Leishmania (family Trypanosomidae). Visceral leishmaniasis (kala-azar), caused by L. donovanni – L. infantum species complex, is of interest to the haematologist. Visceral leishmaniasis (VL) occurs in the Mediterranean littorals, the Balkans, the Middle East, Sudan, East Africa, the Indian subcontinent, China, and South America (‘L. chagasi’). Of the two forms of the Leishmania organism, only the amastigote form (the form that lacks an exteriorized flagellum) is seen in humans in vivo. The promastigote form of the parasite is seen in infected sandflies and after inoculation of aspirated material (e.g. bone marrow) into a special blood agar culture medium called Novy-Nicolle-McNeal (NNN) medium and incubation at 28°C for one to four weeks (Figure 17).

Figure 17. The two forms of Leishmania.
Although amastigotes can occasionally be detected in mononuclear leukocytes or extracellularly in peripheral blood films, the prime means of diagnosis is the detection of amastigotes in bone marrow, lymph node, or splenic aspirates.
Leishmania amastigotes are seen within the phagocytes of the reticuloendothelial system (bone marrow, lymph nodes, spleen, and liver)
Miscoscopical diagnosis of visceral leishmaniasis in bone marrow aspirates
Amastigotes (Leishman-Donovan bodies) are recognised by their characteristic basic structure: they appear as oval or round structures, measuring 2-4 μm in length. Each amastigote has a dark nucleus and a characteristic perinuclear rod-shaped or dot-shaped organelle called kinetoplast within a pale bluish-reddish cytoplasm. The kinetoplast stains red purple.
Note: the morphology of amastigotes is very characteristic. Focus up and down carefully to see the kinetoplast before you identify any structure as a leishmania parasite. To ensure that the observed structures are amastigotes, the kinetoplast must be visualised. If you are uncertain, look for more typical organisms. If you do not find things that are clearly leishmania parasites, report the film as ῾No leishmania amastigotes found᾿.
Amastigotes are typically found inside macrophages (rarely inside neutrophils or metamyelocytes, too). Isolated extracellular amastigotes are also commonly seen in aspirates (Figure 18).

Figure 18. Leishmania amastigotes in a bone marrow aspiration smear.
Bone marrow changes that may be seen in leishmaniasis:
– red cell agglutination and rouleaux
– hypercellularity
– erythroid hyperplasia
– reduced myeloid:erythroid ratio
– left shifted granulopoiesis
– reactive plasmacytosis (often ≥5%) (not in patients taking corticosteroids)
– increased macrophages and histiocytes
– haemophagocytosis (haemophagocytosis is common; development of haemophagocytic syndrome is rare)
– Mott cells (degenerated plasma cells that can be seen in cases of immune system activation by various stimuli), plasma cells with crystals or Russell bodies
– dyserythropoiesis (may be pronounced). Dyserythropoiesis leads to ineffective erythropoiesis.
– megaloblastic change (due to folic acid deficiency)*
– anaemia of chronic disorder (reticuloendothelial iron blockade)
– an increase in lymphocytes (usually mild)
– a mild increase in eosinophil precursors (15% of cases)
– reticulin fibrosis (as a result of chronic bone marrow infection) resulting in a dry tap
– granulomatous reaction with non-caseating granulomas (may be marked)
* In India, 50% of patients with kala-azar have moderate or severe megaloblastic change which responds to folic acid. The severity of megaloblastic change depends on the adequacy of folic acid status and intake of folic acid (diet).
Figure 19 shows that there are four types of bone marrow reaction to chronic infections. In VL, the bone marrow is usually hypercellular or normocellular, with erythropoietic and granulopoietic activity. In longstanding, chronic cases of VL, bone marrow may become fibrotic or even hypoplastic.

Figure 19. The four types of bone marrow reaction to chronic infections.
Note: the FBC consistently shows neutropenia in VL. Treatment is followed by shrinkage of spleen and resolution of pancytopenia but the return of haematological parameters and bone marrow appearances to normality may take 9 months following cure. The haemoglobin response may be delayed due to the anaemia of chronic disorder.
– Amastigotes (intracellular or extracellular) may be demonstrated in:
– bone marrow aspirate (54-86% positivity)
– lymph node aspirate (64% positivity)
– splenic aspirate (98% positivity)
– blood films and buffy coat films made by the microhaematocrit method (monocytes are concentrated in the buffy coat layer): they are occasionally positive in immunocompromised patients e.g. HIV, transplantation, or infants with VL
The parasite density in aspirates of bone marrow, lymph node or spleen is graded using a 100× (oil immersion) objective as follows:
Grade: Average parasite density:
6+ = >100 parasites/field
5+ = 10-100 parasites/field
4+ = 1-10 parasites/field
3+ = 1-10 parasites/10 fields
2+ = 1-10 parasites/100 fields
1+ = 1-10 parasites/1000 fields (These cases are very difficult. Careful examination and a high index of suspicion are required).
Differential diagnosis of leishmania:
Platelets, phagocytosed material, haemosiderin granules, cellular debris, dust, unidentified objects or artefacts may be mistaken for leishmania. These objects however, do not have the basic structure of amastigotes (nucleus-kinetoplast).
T. cruzi amastigotes may rarely be found in bone marrow aspirates of immunocompromised patients from Central or South America as a result of Chagas’ disease reactivation (T. cruzi amastigotes have a larger and more rounded kinetoplast compared to leishmania amastigotes). In such cases, trypomastigote forms may also be found in bone marrow or peripheral bood films in addition to amastigotes.
Figure 20. Morphological characteristics of H. capsulatum.
Leishmania amastigotes must be distinguished from Histoplasma capsulatum, Penicillium marneffei, and Toxoplasma gondii.
Histoplasmosis
Histoplasmosis is caused by the dimorhic fungus H. capsulatum; there are similarities between kala-azar and disseminated histoplasmosis. H. capsulatum enters the phagocytes of the reticuloendothelial system: in disseminated histoplasmosis it is seen inside the macrophages of bone marrow, spleen, lymph nodes, lungs, and liver. ‘Dimorhic fungus’ means that:
– In nature or in culture: H. capsulatum is in mold form.
– In humans: H. capsulatum cells are intracytoplasmic yeasts. They are seen as oval or round inclusions (2-4 μm) with a dot-like or crescent-shaped magenda-coloured cap of chromatin-protoplasm often at one end of the cell and a small capsule (thinner than the capsule of cryptococcus). Some organisms contain an intracytoplasmic vacuole (Figure 20).
The capsule may or may not be visible as a translucent halo surrounding the cell (in fact, this represents retraction of the yeast endoplasm from a very thin cell wall rather than an actual capsule, however, the name H. capsulatum has persevered).
Histoplasma differs from leishmania amastigotes:
– histoplasma is pleomorphic
– histoplasma is found mainly intracellularly
– has a small capsule (not always visible)
– occasional budding forms seen
– absence of kinetoplast
– stains with fungal stains (PAS, Gomori/Grocott) in bone marrow sections
Disseminated histoplasmosis presents with fever, pancytopenia, hepatosplenomegaly, lymphadenopathy, and lung nodules or infiltrates. Disseminated histoplasmosis is more likely to be seen in immunocompromised patients. The bone marrow is typically normocellular normoblastic with numerous macrophages and is frequently found to contain Langhans’ giant cells and granulomas (ddx: granulomatous diseases e.g. tuberculosis). H. capsulatum may also be seen in the peripheral blood film as oval inclusion within a monocyte or neutrophil and should be distinguished from other fungi (e.g. Candida, Cryptococcus, Malassezia, and Penicillium) and leishmania. Histoplasmosis is a cosmopolitan disease: USA (endemic spots in the midwestern states along Mississippi and Ohio rivers and in the southeastern states), Central and South America, Middle East (Arabian Peninsula, Persian Gulf), and Asia (Pakistan, India, China). It is found in soil.
Histoplasmosis is uncommon in South-East Asia. Yet, Penicillium (Talaromyces) marneffei, another dimorphic fungus, causes an illness in South-East Asia and South China which may be mistaken for histoplasmosis or leishmaniasis.
Penicilliosis (talaromycosis)9,15
Disseminated penicilliosis is most likely to be seen in immunocompromised people, particularly those with HIV/AIDS, presenting with fever, pancytopenia, rash, lymphadenopathy, and hepatosplenomegaly.
P. marneffei is found both intracellularly and extracellularly in the bone marrow. The bone marrow aspirates typically show normocellularity with active erythropoiesis and granulopoiesis and many macrophages which contain round, oval, elongated or sausage-shaped inclusions (yeast forms) 2-6 μm with red-purple dot-like nucleus and sometimes transverse septa. Fungal stains are (+). The blood film frequently shows monocytes with ingested fungi.
Microscopical characteristics of P. marneffei:
– septate forms
– elongated or sausage-shaped forms
– extracellular organisms commonly seen (≠ H. capsulatum)
– absence of budding (≠ H. capsulatum)
– absence of kinetoplast (≠ leishmania amastigotes)
Penicilliosis should be considered as a possible explanation for fever and cytopenia in immunocompromised patients from Asia and in those with a history of travel in South-East Asia or South China (Figure 21).

Figure 21. Endemic regions of P. marneffei in Asia.
Toxoplasmosis6,7,16
Toxoplasma gondii tachyzoites may resemble leishmania amastigotes. Tachyzoites (dividing forms) are distinguished by their characteristic bow or crescent shapes (3-6 μm), pointed ends, a small single nucleus, and absence of kinetoplast. Tachyzoites may be found either intracellularly or extracellularly (free) in bone marrow aspirates (Figure 22 A-C); cystic forms (pseudocysts or cysts containing numerous bradyzoites) may also be present. Toxoplasma has also been identified in the peripheral blood, either extracellularly or within neutrophils (Figure 22 D). Neither septa nor yeast forms occur in T. gondii. Gomori’s stain is negative. Immunocompromised hosts with disseminated toxoplasmosis present with fever, pancytopenia, hepatomegaly, and interstitial pneumonia, with or without CNS lesions.
Remember:
Leishmania
Histoplasma
Toxoplasma
Penicillium
may be found in bone marrow and blood films of immunocompromised patients (ΗIV, haematological malignancy, transplantation, corticosteroids) with pancytopenia and fever (look for the presence or absence of kinetoplast).
1.5 TROPICAL BACTERIAL INFECTIONS THAT CAN BE DIAGNOSED FROM THE BLOOD FILM66-12
The blood film may demostate toxic neutrophils (vacuoles and toxic granulation) with phagocytosed bacteria in hyposplenic or immunocompromised patients and in those with central venous lines or overwhelming septicaemia. Bacteria that have been phagocytosed by neutrophils appear as blue inclusions in Giemsa e.g. Staphylococcus spp., S. pneumoniae, C. perfringens, Ν. meningitis, Ehrlichia spp., and Capnocytophaga canimorsus. Of these, infection with Νeisseria meningitis type A occurs in the tropics.

Figure 22. A-D. Disseminated toxoplasmosis.
Note: if the organisms are intracellular, it is definite evidence of a bacteraemia or septicaemia. If the organisms are only extracellular, then a source of contamination in the environment must first be ruled out.
The ‘meningitis belt’ of tropical Africa is the zone lying between 5ο and 15ο N of the Equator, and characterised by an annual rainfall between 30 and 110 cm (Figure 23). In the ‘meningitis belt’ of Africa, type Α Νeisseria meningitis is the usual causative agent of epidemics. N. meningitis is a Gram (-), coffee bean shaped diplococcal organism (Figure 24).

Figure 23. The meningitis belt of Africa.

Figure 24. A 36-year-old woman with rash (irregular scattered petechiae), headache, and fever. Meningococcaemia (phagocytosed diplococci).
Two arthropod-borne tropical bacterial infections are diagnosed from the blood film:
1. The relapsing fevers: two forms of relapsing fever exist:
– tick-borne relapsing fever caused by about 15 Borrelia species, especially Borrelia duttoni. Soft ticks such as Ornithodoros spp. transmit B. duttoni from rodents (mice) to man. First described from Africa (B. duttoni is endemic in East Africa e.g. Τanzania), tick-borne relapsing fever also occurs in USA (forests of western and north-western states where it is frequently due to Β. hermsii), the Middle East, and the Mediterranean basin (especially Spain and North Africa). It has been described in Greece. Infection is more common in children.
– louse-borne relapsing fever (caused by Borrelia recurrentis). Borrelia recurrentis, transmitted by body lice, occurs in epidemic form only in Ethiopia, Eritrea, and South Sudan. Mortality is high.
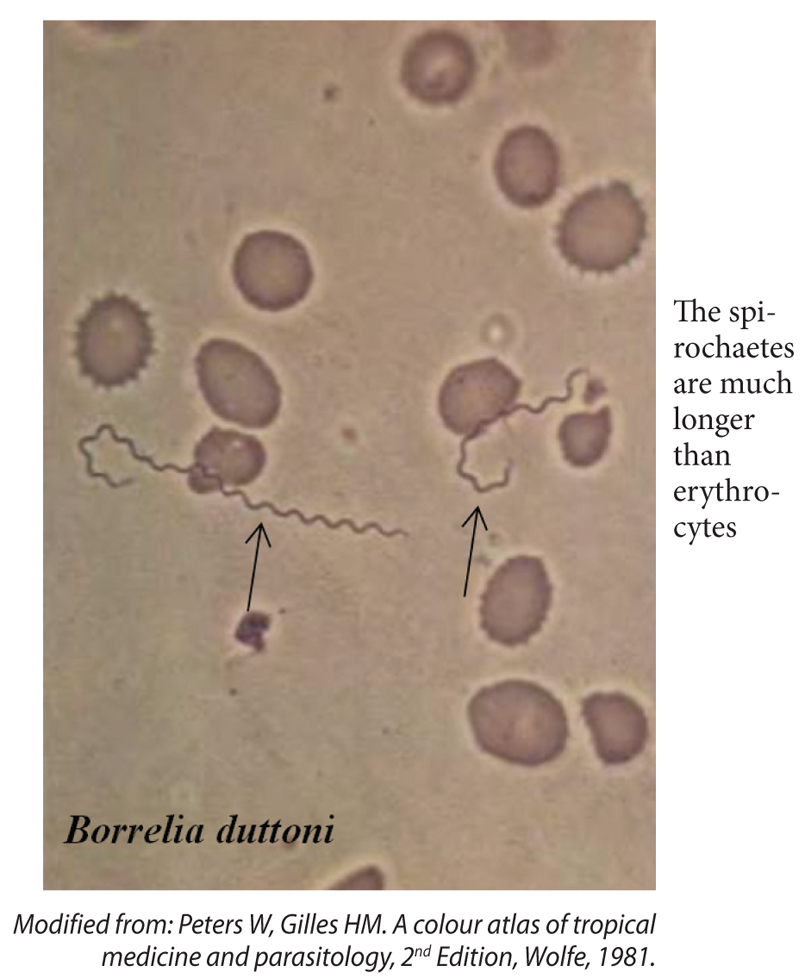
Figure 25. B. duttoni in a peripheral blood film.
The disease acquired its name from the typical relapsing nature of the fever. The febrile periods recur on one or two occasions only. Laboratory tests show transaminitis (ddx: viral hepatitis), leukocytosis (e.g. neutrophils 90%), and thrombocytopenia. Leukopenia is uncommon (unlike malaria). Tick-borne relapsing fever is usually milder than the louse-borne form. Infection is more serious in pregnant women. Neurological involvement (meningoencephalitis) because may occur because borrelias are neurotropic.
Note: relapsing fever may clinically resemble malaria.
Relapsing fever may be diagnosed in travellers and migrants returning from Africa with a malaria like picture (intermittent fever, chills, headache, and splenomegaly).
Borrelia spirochaetes only appear in the blood during the febrile episodes. They are numerous! They are seen in thin films between the red cells (Figure 25) or in thick films. Borrelia recurrentis and B. duttoni are morphologically indistinguishable. They are long spiral organisms (15-30 μm × 0.4 μm) with 5-10 (often 6-8) fairly regular but loose waves (corkscrew appearance) which stain pink or deep pink in Giemsa. They have tapering ends. They multiply in blood by transverse fission. The organisms are also easily seen in India ink preparations. B. duttoni should not be confused with the exflagellated microgametocyte of malaria parasites.
The incubation period of relapsing fever is 4-8 days after exposure (range: 2-15 days)
2. Bartonellosis (Carrión’s disease or Οroya fever) caused by Bartonella bacilliformis, a small pleomorphic gram-negative intraerythrocytic bacterium (coccobacillus) which produces fever and acute haemolytic anaemia in Peru, Bolivia, Colombia, and Ecuador (anaemic phase). The bacilli also invade reticulo-endothelial cells. About 40% of those with untreated bartonellosis die in the anaemic phase. Lymphadenopathy and splenomegaly may be present due to erythrophagocytosis. During recovery from the anaemic phase one or two months later, a generalised verrucous eruption angiomatous in nature may develop, known as Verruga Peruviana (eruptive phase). Individuals with a history of Oroya fever and those with Verruga Peruviana must be excuded from blood donation because they can transmit the disease (blood cultures are positive). More than 15% of the residents of the Peruvian Andes are latently infected. Therefore, people originating from this region, even without a history of infection, should be excluded from blood donation. Like malaria and babesiosis, bartonellosis is particularly dangerous for asplenic patients.
The infection is transmitted by the sandfly Lutzomyia verrucarum at altitudes of 800-3000 m among those who live in, or those mountaineering in, the Andes in Peru, Colombia, or Ecuador (Figure 26). However, most infections are asymptomatic or oligosymptomatic.

Figure 26. Established endemic foci of bartonellosis are found in Peru, Ecuador, and Colombia.
The clinical picture of Carrión’s disease (which lasts 2-3 weeks) is that of a febrile Coombs (-) haemolytic anaemia, often macrocytic. The blood film demonstrates numerous Bartonella organisms within erythrocytes (Figure 27). The organisms seen in blood are coccoid to rod-like, sometimes bayonet-shaped, curved or bipolar-stained coccobacilli (markedly pleomorphic coccobacilli). They are small, measuring 1-3 × 0.25-0.5 μm. They often form chains of three and may be seen as V or Y forms. The organisms stain red, deep red or purple with Giemsa; they do not stain with Gram’s stain. In severe infections, 90-100% of erythrocytes may contain bacilli and up to 12 organisms can be found per rbc. Circulating erythrocytes in a few days may fall from normal to less than 1 million. Haemolytic anaemia is due to intravascular haemolysis with gross haemoglobinuria and due to splenic removal of deformed infected erythrocytes. Microscopical findings include striking spherocytosis (spherocytes, microsperocytes), polychromasia and prominent fine basophilic stippling, nucleated red cells, crenated cells, stomatocytes, red cells with an irregular outline, and decoloured (hypochromic) red cells. There are no blister cells. The leukocyte count is increased with neutrophilia (neutrophils 90%) and toxic granulation (because bartonellosis is a bacterial infection), but neutrophils with phagocytosed organisms are not seen. Bartonellosis may be complicated by secondary septicaemia, commonly caused by salmonellae (typhoid fever).

Figure 27. Blood film of a patient with Carrión’s disease.
Note: a mixture of coccobacilli is seen:
– more common in the early days are thin, rod-like bacillary forms 1-3 μm in diametre
– more common after a few days are red-purple coccoid forms about 0.5 μm in diametre
Blood culture of B. bacilliformis is difficult: B. bacilliformis is a fastidious bacterium and culture requires a specific medium (Columbia agar) incubated at 25-28°C for 1-2 weeks. In practice, diagnosis of bartonellosis depends upon microscopy.
The incubation period of Carrión’s disease is 3-8 weeks (up to 9 months in some cases).
Tips:
– when taking a medical history, questions about travel are of paramount importance. Bartonellosis should be suspected in tourists returning from Peru with fever and anaemia. The most important ddx is malaria. P. vivax is the only species of Plasmodium prevalent in the highlands of Peru (Babesia is not prevalent in South America).
– Bartonella organisms should be distinguished from rbc staining artefacts (e.g. precipitated stain on red blood cells, unfiltered Giemsa, dust, debris from the patient’s skin etc).
– other bartonellae i.e. B. henselae (cat scratch disease) and B. quintana (bacillary angiomatosis, endocarditis, trench fever) which is transmitted by body lice are not detected in blood films.
– T. whippeli bacilli causing fever and haemolytic anaemia may be seen in the blood film as numerous delicate intraerythrocytic delicate rod-shaped bacilli in hyposplenic patients with Whipple’s disease which may resemble bartonellae (T. whippeli is haemotropic).
1.6 ILLUSTRATIONS OF BLOOD PARASITES
Malaria (Figures 28-38)
Figure 28. A-F. A 24-year-old man with intermittent fever who had visited his parents in Nigeria. He was an international student at the University of Crete. The figures show P. falciparum rings and gametocytes.

Figure 29. A and B. Α 50-year-old patient with severe malaria, haemoglobinuria, jaundice, and renal failure. P. falciparum rings and schizonts.

Figure 30. A and B. P. falciparum infection showing late trophozoites, Maurer’s dots, and rings.

Figure 31. A-D. Extreme P. falciparum parasitaemia (35%) with numerous small rings (ddx: B. microti coccoid forms) in a 45-year-old woman who presented with febrile haemolytic anaemia. Note the presence of toxic neutrophils in panel C and the banana-shaped female gametocyte in panel D.

Figure 32. A and B. A 45-year-old migrant from Cameroon with fever 39°C, productive cough, and generalised weakness. Only gametocytes of P. falciparum were seen in the blood film.

Figure 33. A-D. A 42-year-old man, originally from Pakistan, who has been is living for 10 years in Crete and has not travelled outside Greece for 5 years. He presented with fever and myalgia of 3 days’ duration. chüffner’s dots, amoeboid trophozoites, and rings in enlarged red cells consistent with P. vivax relapse.

Figure 34. A-F. A 42-year-old woman who returned from a 2-month trip to India 2 days ago and is now not feeling well. Blood tests showed WBC 5.8 ×109/l, Hb 12.8 g/dl, PLT 75×109/l, MCV 87.5 fl. Schüffner’s dots, amoeboid trophozoites, gametocytes, and ring forms consistent with P. vivax malaria.

Figure 35. A-F. A 30-year-old migrant from Afghanistan with fever 40°C and headache. Blood tests showed WBC 3.9×109/l, Hb 14.6 g/dl, PLT 113×109/l, MPV 11.0 fl, MCV 93 fl. P. falciparum RDT was negative. P. vivax rings, amoeboid trophozoites, schizonts, free merozoites, and ookinete.

Figure 36. A-F. A 4-year-old girl who visited her grandmother in Ghana. After returning to London, she was diagnosed with P. οvale malaria. James’ dots, trophozoites, and gametocytes.

Figure 37. A and B. A 35-year-old Ghanaian woman with myalgia and arthralgia for the past 4 weeks. P. malariae trophozoites and schizonts.

Figure 38. Malaria parasites and other red cell disorders. Panel A. Iron deficiency anaemia and P. falciparum. Panel B. Heterozygous β-thalassaemia and P. falciparum. Panel C. Heterozygous HbC and P. malariae. Panel D. African elliptocytosis and P. falciparum. Panels E and F. Sickle cell anaemia and P. ovale.
African trypanosomiasis (Figures 39 and 40)

Figure 39. A-B. A 55-year-old man who returned from a safari in East Africa 6 days ago. Fevers, reduced consciousness, diarrhoea, and tachycardia (150 bpm). Thick film with Τ. b. rhodesiense trypomastigotes.

Figure 40. A-F. A 29-year-old Greek sailor with a history of journeys to West Africa (Senegal, Gambia, and Ghana) and Cameroon presented with cervical and inguinal lymphadenopathy. Afebrile, spleen tip just palpable. History of malaria 5 years earlier. WBC normal, ESR normal. Rare Τ. b. gambiense trypomastigotes.
Μicrofilariae (Figures 41-44)
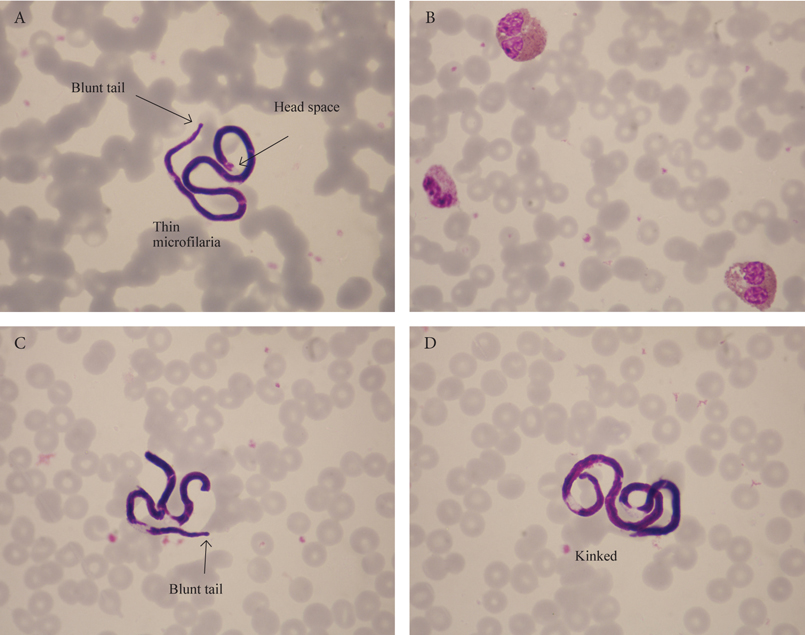
Figure 41. A-D. A 23-year-old woman who has recently returned from her honeymoon in Egypt and is not ῾feeling herself᾿ (difficulty in concentration, nausea). Blood tests showed eosinophilia. Daytime capillary blood film showing Mansonella perstans microfilariae.

Figure 42. A-D. A 67-year-old woman noted pruritus, swelling, and mild pain of her left wrist for 10 days, without antecedent trauma. 8 months earlier she had visited her brother who works in Cameroon. Blood tests showed eosinophilia. Daytime capillary thin blood film showing Loa loa infection.

Figure 43. A-D. A 50-year-old man from India with eosinophilia and lymphadenopathy. Lymph node biopsy showed marked infiltration with eosinophils and eosinophilic abscesses. Night thick blood film showing Wuchereria bancrofti.

Figure 44. A-D. Night capillary thin blood film from a patient from Cambodia with lymphoedema and a normal eosinophil count. Brugia malayi lymphatic filariasis.
Leishmaniasis (Figures 45-48)

Figure 45. A and B. A 58-year-old Greek woman with pancytopenia, fever, and hepatosplenomegaly. Bone marrow findings were typical of kala-azar.

Figure 46. A and B. A 23-year-old Greek man who had undergone allogeneic stem cell transplant for AML. Presented with fever and pancytopenia. He had no splenomegaly. Visceral leishmaniasis in immunocompromised host.

Figure 47. A and B. A 75-year-old Greek woman with rheumatoid arthritis, fever of one weeks’ duration, leukopenia (PMN 0.7×109/l), anaemia (Hb 11 g/dl), PLT 300×109/l, and splenomegaly. Bone marrow aspiration was a dry tap because of reticulin fibrosis. Trephine rolls showed scattered amastigotes.

Faced with a difficult diagnosis or uncertainty, haematologists should consult with morphologists who have expertise in the field of microscopical diagnosis of tropical diseases.
I have found the following key references of considerable value in preparing this manuscript. Many further references will be found in each of these works.
References
- World Health Organisation. Βench aids for the diagnosis of malaria. Geneva: WHO; c1988.
- Warhurst DC, Williams JE. ACP Broadsheet no 148. July 1996. Laboratory diagnosis of malaria. J Clin Pathol. 1996 Jul;49(7):533-8.
- Mackie TΤ, Hunter GW III, Brooke Worth C. A manual of tropical medicine. Philadelphia: WB Saunders Company; c1945.
- World Health Organisation. Βasic laboratory methods in medical parasitology. Geneva: WHO; c1991.
- Walter Reed Army Institute of Research (WRAIR) Tropical Medicine Course, Laboratory Manual. 5th ed. Silver Springs: Walter Reed Army Institute of Research; c2013.
- Peters W, Gilles HM. A colour atlas of tropical medicine and parasitology. 2nd ed. London: Wolfe; c1981.
- Peters W, Pasvol G. A colour atlas of tropical medicine and parasitology. 6th ed. London: Mosby Elsevier; c2007.
- Cook GC, Zumla AI. Manson’s tropical disease. 22nd ed. London: Εlsevier Saunders; c2008.
- Farrar J, Hotez P, Junghanss T, Kang G, Lalloo D, White N. Manson’s tropical diseases. 23rd ed. London: Εlsevier Saunders; c2014.
- Binford CH, Connor DH. Pathology of tropical and extraordinary diseases. Washington: Armed Forces Institute of Pathology; c1976.
- Bain BJ. Blood cells – a practical guide. 3rd ed. London: Blackwell Publishing; c2002.
- Kapff C, Jandl J. Blood atlas and sourcebook of hematology. 2nd ed. Boston: Little Brown and Company; c1991.
- World Health Organisation. Βench aids for the diagnosis of filarial infections. Geneva: WΗO; c1997.
- Liapis K, Chung Y, Akhtar W, Taussig D. A patient with rheumatoid arthritis, cryoglobulinaemia, and an “accidental” finding. BMJ. 2014 Apr;348:g2701.
- Mandell GL, Diamond RD. Atlas of infectious diseases. Fungal infections. Philadelphia: Current Medicine; c2000.
- Liapis K, Baltadakis I, Balotis K, Karakasis D. Multisystem febrile illness in a cord-blood transplant recipient. Clin Infect Dis. 2010 Sep;51(6):716-7.
- Spira AM. Assessment of travellers who return home ill. Lancet. 2003 Apr;361(9367):1459-69.
- Ryan ET, Wilson ME, Kain KC. Illness after international travel. N Engl J Med. 2002 Aug;347(7):505-16.